


















































 Islands")


















































")






")
















































")
























































































")
")





 Germany
Germany
 Japan
Japan
 United Kingdom
United Kingdom
 China
China
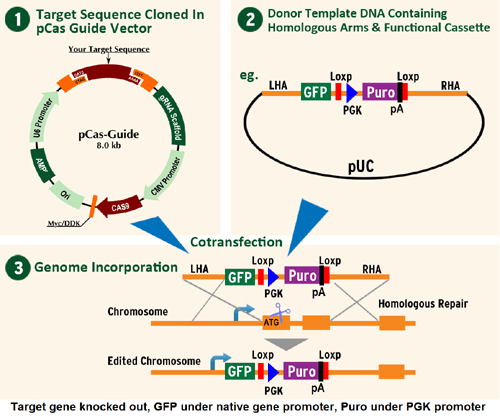
Ddx3x Mouse Gene Knockout Kit (CRISPR)
CAT#: KN304393
Ddx3x - mouse gene knockout kit via CRISPR, HDR mediated
Functional Cassette: Luciferase-Puro RFP-BSD mBFP-Neo
HDR-mediated knockout kit validation
USD 1,657.00
2 Weeks*
Product Images
Specifications
| Product Data | |
| Format | 2 gRNA vectors, 1 GFP-puro donor, 1 scramble control |
| Donor DNA | GFP-puro |
| Symbol | Ddx3x |
| Locus ID | 13205 |
| Components |
KN304393G1, Ddx3x gRNA vector 1 in pCas-Guide CRISPR vector, Target Sequence: AAGTGATGCAGCGACCCCCG KN304393G2, Ddx3x gRNA vector 2 in pCas-Guide CRISPR vector, Target Sequence: GCAGGTGAGCCTGAGCCGCG KN304393D, donor DNA containing left and right homologous arms and GFP-puro functional cassette. Homologous arm and GFP-puro sequences: pUC vector backbone in gray; Left arm sequence in blue; GFP-puro in green; Right arm in violet GATCGTTGGG AACCGGAGCT GAATGAAGCC ATACCAAACG ACGAGCGTGA CACCACGATG CCTGTAGCAA TGGCAACAAC GTTGCGCAAA CTATTAACTG GCGAACTACT TACTCTAGCT TCCCGGCAAC AATTAATAGA CTGGATGGAG GCGGATAAAG TTGCAGGACC ACTTCTGCGC TCGGCCCTTC CGGCTGGCTG GTTTATTGCT GATAAATCTG GAGCCGGTGA GCGTGGTTCT CGCGGTATCA TTGCAGCACT GGGGCCAGAT GGTAAGCCCT CCCGTATCGT AGTTATCTAC ACGACGGGGA GTCAGGCAAC TATGGATGAA CGAAATAGAC AGATCGCTGA GATAGGTGCC TCACTGATTA AGCATTGGTA ACTGTCAGAC CAAGTTTACT CATATATACT TTAGATTGAT TTAAAACTTC ATTTTTAATT TAAAAGGATC TAGGTGAAGA TCCTTTTTGA TAATCTCATG ACCAAAATCC CTTAACGTGA GTTTTCGTTC CACTGAGCGT CAGACCCCGT AGAAAAGATC AAAGGATCTT CTTGAGATCC TTTTTTTCTG CGCGTAATCT GCTGCTTGCA AACAAAAAAA CCACCGCTAC CAGCGGTGGT TTGTTTGCCG GATCAAGAGC TACCAACTCT TTTTCCGAAG GTAACTGGCT TCAGCAGAGC GCAGATACCA AATACTGTTC TTCTAGTGTA GCCGTAGTTA GGCCACCACT TCAAGAACTC TGTAGCACCG CCTACATACC TCGCTCTGCT AATCCTGTTA CCAGTGGCTG CTGCCAGTGG CGATAAGTCG TGTCTTACCG GGTTGGACTC AAGACGATAG TTACCGGATA AGGCGCAGCG GTCGGGCTGA ACGGGGGGTT CGTGCACACA GCCCAGCTTG GAGCGAACGA CCTACACCGA ACTGAGATAC CTACAGCGTG AGCTATGAGA AAGCGCCACG CTTCCCGAAG GGAGAAAGGC GGACAGGTAT CCGGTAAGCG GCAGGGTCGG AACAGGAGAG CGCACGAGGG AGCTTCCAGG GGGAAACGCC TGGTATCTTT ATAGTCCTGT CGGGTTTCGC CACCTCTGAC TTGAGCGTCG ATTTTTGTGA TGCTCGTCAG GGGGGCGGAG CCTATGGAAA AACGCCAGCA ACGCGGCCTT TTTACGGTTC CTGGCCTTTT GCTGGCCTTT TGCTCACATG TTCTTTCCTG CGTTATCCCC TGATTCTGTG GATAACCGTA TTACCGCCTT TGAGTGAGCT GATACCGCTC GCCGCAGCCG AACGACCGAG CGCAGCGAGT CAGTGAGCGA GGAAGCGGAA GAGCGCCCAA TACGCAAACC GCCTCTCCCC GCGCGTTGGC CGATTCATTA ATGCAGCTGG CACGACAGGT TTCCCGACTG GAAAGCGGGC AGTGAGCGCA ACGCAATTAA TGTGAGTTAG CTCACTCATT AGGCACCCCA GGCTTTACAC TTTATGCTTC CGGCTCGTAT GTTGTGTGGA ATTGTGAGCG GATAACAATT TCACACAGGA AACAGCTATG ACCATGATTA CGCCAAGCTC CTTCCTCTTC CAGCCCTTCC TCTTTCACAC TGACTGACTG ACCTGCATAC ACCTCCCACG GTCACGTGAT CCCACTGCCC TCCACGCAGA GCCAGGGCTC ATTAGTTAGG CCAGACAAAG CCCACTGCAT CATCTCCACG CCCGGGGACG GAGAGGAAGG GACGAGTGCT AGAAAGAAGG ACGAAAACTG GGTACACAGG GGAGGAGCAG CCAATCAACC TAGTTTAAAG CTTAGGGCCT TAAGGCTAGA GGACGGTCAG CAATCATTTC TCGCGAGATC TCGAGAAAAC AGTAAGATTA GGGGACCGGT GGAGACCAGG GGATAGCAAA CCCAGGAGAA CCAGAACCAC ATATACCCAA GGCTTCTTTA TGAGCCGACG AGGACGCGGG TAACTGGACT GATTAGGAAA TCTCACGGGA CTCAGCCGAG CAACGCACCG TAAGCAGGCA GAGCTCCTAA CGGCAGTGGC CGGCCGCCCG GGAGAGGAGG GCACACGTCT TACGTGCTGA CGTAGCAGTC CTCCGAGTCG GTATATAAGG TCGGTGGCCG CGCGGTGCGC TTCAAAGCCG CAGTTCTCCC GTGAGAGGGC CTTTGCGGTA GAGGCAGCCT TTGCTCAGCA GCGGAAGACT CCGATTTCTC GGTACTCTTC AGGGACTAGC ATGGAGAGCG ACGAGAGCGG CCTGCCCGCC ATGGAGATCG AGTGCCGCAT CACCGGCACC CTGAACGGCG TGGAGTTCGA GCTGGTGGGC GGCGGAGAGG GCACCCCCGA GCAGGGCCGC ATGACCAACA AGATGAAGAG CACCAAAGGC GCCCTGACCT TCAGCCCCTA CCTGCTGAGC CACGTGATGG GCTACGGCTT CTACCACTTC GGCACCTACC CCAGCGGCTA CGAGAACCCC TTCCTGCACG CCATCAACAA CGGCGGCTAC ACCAACACCC GCATCGAGAA GTACGAGGAC GGCGGCGTGC TGCACGTGAG CTTCAGCTAC CGCTACGAGG CCGGCCGCGT GATCGGCGAC TTCAAGGTGA TGGGCACCGG CTTCCCCGAG GACAGCGTGA TCTTCACCGA CAAGATCATC CGCAGCAACG CCACCGTGGA GCACCTGCAC CCCATGGGCG ATAACGATCT GGATGGCAGC TTCACCCGCA CCTTCAGCCT GCGCGACGGC GGCTACTACA GCTCCGTGGT GGACAGCCAC ATGCACTTCA AGAGCGCCAT CCACCCCAGC ATCCTGCAGA ACGGGGGCCC CATGTTCGCC TTCCGCCGCG TGGAGGAGGA TCACAGCAAC ACCGAGCTGG GCATCGTGGA GTACCAGCAC GCCTTCAAGA CCCCGGATGC AGATGCCGGT GAAGAAAGAG TTTAAGAATT CCGATCATAT TCAATAACCC TTAATATAAC TTCGTATAAT GTATGCTATA CGAAGTTATT AGGTCTGAAG AGGAGTTTAC GTCCAGCCAA GCTTAGGATC TCGACCTCGA AATTCTACCG GGTAGGGGAG GCGCTTTTCC CAAGGCAGTC TGGAGCATGC GCTTTAGCAG CCCCGCTGGG CACTTGGCGC TACACAAGTG GCCTCTGGCC TCGCACACAT TCCACATCCA CCGGTAGGCG CCAACCGACT CCGTTCTTTG GTGGCCCCTT CGCGCCACCT TCTACTCCTC CCCTAGTCAG GAAGTTCCCC CCCGCCCCGC AGCTCGCGTC GTGCAGGACG TGACAAATGG AAGTAGCACG TCTCACTAGT CTCGTGCAGA TGGACAGCAC CGCTGAGCAA TGGAAGCGGG TAGGCCTTTG GGGCAGCGGC CAATAGCAGC TTTGCTCCTT CGCTTTCTGG GCTCAGAGGC TGGGAAGGGG TGGGTCCGGG GGCGGGCTCA GGGGCGGGCT CAGGGGCGGG GCGGGCGCCC GAAGGTCCTC CGGAGGCCCG GCATTCTGCA CGCTTCAAAA GCGCACGTCT GCCGCGCTGT TCTCCTCTTC CTCATCTCCG GGCCTTTCGA CCTGCATCCA TCTAGATCTC GAGCAGCTGA AGCTTACCAT GACCGAGTAC AAGCCCACGG TGCGCCTCGC CACCCGCGAC GACGTCCCCA GGGCCGTACG CACCCTCGCC GCCGCGTTCG CCGACTACCC CGCCACGCGC CACACCGTCG ATCCGGACCG CCACATCGAG CGGGTCACCG AGCTGCAAGA ACTCTTCCTC ACGCGCGTCG GGCTCGACAT CGGCAAGGTG TGGGTCGCGG ACGACGGCGC CGCGGTGGCG GTCTGGACCA CGCCGGAGAG CGTCGAAGCG GGGGCGGTGT TCGCCGAGAT CGGCCCGCGC ATGGCCGAGT TGAGCGGTTC CCGGCTGGCC GCGCAGCAAC AGATGGAAGG CCTCCTGGCG CCGCACCGGC CCAAGGAGCC CGCGTGGTTC CTGGCCACCG TCGGCGTCTC GCCCGACCAC CAGGGCAAGG GTCTGGGCAG CGCCGTCGTG CTCCCCGGAG TGGAGGCGGC CGAGCGCGCC GGGGTGCCCG CCTTCCTGGA GACCTCCGCG CCCCACAACC TCCCCTTCTA CGAGCGGCTC GGCTTCACCG TCACCGCCGA CGTCGAGGTG CCCGAAGGAC CGCGCACCTG GTGCATGACC CGCAAGCCCG GTGCCTGACG CCCGCCCCAC GACCCGCAGC GCCCGACCGA AAGGAGCGCA CGACCCCATG CATCGATGAT ATCAGATCCC CGGGATGCAG AAATTGATGA TCTATTAAAC AATAAAGATG TCCACTAAAA TGGAAGTTTT TCCTGTCATA CTTTGTTAAG AAGGGTGAGA ACAGAGTACC TACATTTTGA ATGGAAGGAT TGGAGCTACG GGGGTGGGGG TGGGGTGGGA TTAGATAAAT GCCTGCTCTT TACTGAAGGC TCTTTACTAT TGCTTTATGA TAATGTTTCA TAGTTGGATA TCATAATTTA AACAAGCAAA ACCAAATTAA GGGCCAGCTC ATTCCTCCCA CTCATGATCT ATAGATCTAT AGATCTCTCG TGGGATCATT GTTTTTCTCT TGATTCCCAC TTTGTGGTTC TAAGTACTGT GGTTTCCAAA TGTGTCAGTT TCATAGCCTG AAGAACGAGA TCAGCAGCCT CTGTTCCACA TACACTTCAT TCTCAGTATT GTTTTGCCAA GTTCTAATTC CATCAGAAGC TGGTCGAGAT CCGGAACCCT TAATATAACT TCGTATAATG TATGCTATAC GAAGTTATTA GGTCCCTCGA AGAGGTTCAC TAGGCGCGCC TTGTGGCGCA GCTCTTTAAG TTCTGCTTTT TGCCTTCCCG TCTCCCTGGA CGGCTCGGGC CGGCGGGTGG CCAGTGGCTA GGGGCTCTGG AGGGACTGGA TGCTTGCGGG GGGAGTGCTA GAGCCGGTGA TTCCTCCGGC TCGGCTCGGT GCGCGGTATT GTCCCTGGCC CGAGCACAAT GGCGGCTTTT GTGTCTGTGC GCAGGCGAGC GGAGGGGGAG GGGGCGCGCG CATTCGGGCG CACGCGCGTC CCCTGGTTGA TTAGTGACCC GCAGGTGGGG ATCGGGGCGG TCGGGGGGGG GAGATTGCCC CGCGTGGCGT GGCGGGGCCC GGGTGGATGG TTTTGGCCCC GGATTTCCGG GGCTTTGGGC TTCCATGGCC CCAGTGGCTG TGGTTTGGAA ATTAGGGAGC TGGCGGCGGC GCGCGGGCTG GCCGAGTGGC CCCGCGGCGC TGGAATGTGG GAGGGGGCGC CGCGGGAGGG GGGCTGGCCT CTGCGCGGGC TAATGGCGGC GGCCTCCTTT GTGTGGCGCA GGCCGGCGGT GCTGCGCGGT CGCCCCCCCA CCCCCATCCC CATCCCCCCG GGTCTGCCCG CCAGCGGGTT AGCACAGTCT TCACTGACTG ACTGACTGGA AAGTCCTCTC CACTGACTGT AGCCTCCAAT TCACTGGCCG TCGTTTTACA ACGTCGTGAC TGGGAAAACC CTGGCGTTAC CCAACTTAAT CGCCTTGCAG CACATCCCCC TTTCGCCAGC TGGCGTAATA GCGAAGAGGC CCGCACCGAT CGCCCTTCCC AACAGTTGCG CAGCCTGAAT GGCGAATGGC GCCTGATGCG GTATTTTCTC CTTACGCATC TGTGCGGTAT TTCACACCGC ATACGTCAAA GCAACCATAG TACGCGCCCT GTAGCGGCGC ATTAAGCGCG GCGGGTGTGG TGGTTACGCG CAGCGTGACC GCTACACTTG CCAGCGCCCT AGCGCCCGCT CCTTTCGCTT TCTTCCCTTC CTTTCTCGCC ACGTTCGCCG GCTTTCCCCG TCAAGCTCTA AATCGGGGGC TCCCTTTAGG GTTCCGATTT AGTGCTTTAC GGCACCTCGA CCCCAAAAAA CTTGATTTGG GTGATGGTTC ACGTAGTGGG CCATCGCCCT GATAGACGGT TTTTCGCCCT TTGACGTTGG AGTCCACGTT CTTTAATAGT GGACTCTTGT TCCAAACTGG AACAACACTC AACCCTATCT CGGGCTATTC TTTTGATTTA TAAGGGATTT TGCCGATTTC GGCCTATTGG TTAAAAAATG AGCTGATTTA ACAAAAATTT AACGCGAATT TTAACAAAAT ATTAACGTTT ACAATTTTAT GGTGCACTCT CAGTACAATC TGCTCTGATG CCGCATAGTT AAGCCAGCCC CGACACCCGC CAACACCCGC TGACGCGCCC TGACGGGCTT GTCTGCTCCC GGCATCCGCT TACAGACAAG CTGTGACCGT CAACGGGAGC TGCATGTGTC AGAGGTTTTC ACCGTCATCA CCGAAACGCG CGACCCGAAA GGGCCTCGTG ATACGCCTAT TTTTATAGGT TAATGTCATG ATAATAATGG TTTCTTAGAC GTCAGGTGGC ACTTTTCGGG GAAATGTGCG CGGAACCCCT ATTTGTTTAT TTTTCTAAAT ACATTCAAAT ATGTATCCGC TCATGAGACA ATAACCCTGA TAAATGCTTC AATAATATTG AAAAAGGAAG AGTATGAGTA TTCAACATTT CCGTGTCGCC CTTATTCCCT TTTTTGCGGC ATTTTGCCTT CCTGTTTTTG CTCACCCAGA AACGCTGGTG AAAGTAAAAG ATGCTGAAGA TCAGTTGGGT GCACGAGTGG GTTACATCGA ACTGGATCTC AACAGCGGTA AGATCCTTGA GAGTTTTCGC CCCGAAGAAC GTTTTCCAAT GATGAGCACT TTTAAAGTTC TGCTATGTGG CGCGGTATTA TCCCGTATTG ACGCCGGGCA AGAGCAACTC GGTCGCCGCA TACACTATTC TCAGAATGAC TTGGTTGAGT ACTCACCAGT CACAGAAAAG CATCTTACGG ATGGCATGAC AGTAAGAGAA TTATGCAGTG CTGCCATAAC CATGAGTGAT AACACTGCGG CCAACTTACT TCTGACAACG ATCGGAGGAC CGAAGGAGCT AACCGCTTTT TTGCACAACA TGGGGGATCA TGTAACTCGC CTTGE100003, scramble sequence in pCas-Guide vector |
| Disclaimer | These products are manufactured and supplied by OriGene under license from ERS. The kit is designed based on the best knowledge of CRISPR technology. The system has been functionally validated for knocking-in the cassette downstream the native promoter. The efficiency of the knock-out varies due to the nature of the biology and the complexity of the experimental process. |
| Reference Data | |
| RefSeq | NM_010028 |
| UniProt ID | Q62167 |
| Synonyms | D1Pas1-rs2; Ddx3; Fin14 |
| Summary | Multifunctional ATP-dependent RNA helicase. The ATPase activity can be stimulated by various ribo- and deoxynucleic acids indicative for a relaxed substrate specificity. In vitro can unwind partially double-stranded DNA with a preference for 5'-single-stranded DNA overhangs. Is involved in several steps of gene expression, such as transcription, mRNA maturation, mRNA export and translation. However, the exact mechanisms are not known and some functions may be specific for a subset of mRNAs. Involved in transcriptional regulation. Can enhance transcription from the CDKN1A/WAF1 promoter in a SP1-dependent manner. Found associated with the E-cadherin promoter and can down-regulate transcription from the promoter. Involved in regulation of translation initiation. Proposed to be involved in positive regulation of translation such as of cyclin E1/CCNE1 mRNA and specifically of mRNAs containing complex secondary structures in their 5'UTRs; these functions seem to require RNA helicase activity. Specifically promotes translation of a subset of viral and cellular mRNAs carrying a 5'proximal stem-loop structure in their 5'UTRs and cooperates with the eIF4F complex. Proposed to act prior to 43S ribosomal scanning and to locally destabilize these RNA structures to allow recognition of the mRNA cap or loading onto the 40S subunit. After association with 40S ribosomal subunits seems to be involved in the functional assembly of 80S ribosomes; the function seems to cover translation of mRNAs with structured and non-structured 5'UTRs and is independent of RNA helicase activity. Also proposed to inhibit cap-dependent translation by competetive interaction with EIF4E which can block the EIF4E:EIF4G complex formation. Proposed to be involved in stress response and stress granule assembly; the function is independent of RNA helicase activity and seems to involve association with EIF4E. May be involved in nuclear export of specific mRNAs but not in bulk mRNA export via interactions with XPO1 and NXF1. Also associates with polyadenylated mRNAs independently of NXF1. Associates with spliced mRNAs in an exon junction complex (EJC)-dependent manner and seems not to be directly involved in splicing. May be involved in nuclear mRNA export by association with DDX5 and regulating its nuclear location. Involved in innate immune signaling promoting the production of type I interferon (IFN-alpha and IFN-beta); proposed to act as viral RNA sensor, signaling intermediate and transcriptional coactivator. Involved in TBK1 and IKBKE-dependent IRF3 activation leading to IFNB induction, plays a role of scaffolding adapter that links IKBKE and IRF3 and coordinates their activation. Also found associated with IFNB promoters; the function is independent of IRF3. Can bind to viral RNAs and via association with MAVS/IPS1 and DDX58/RIG-I is thought to induce signaling in early stages of infection. Involved in regulation of apoptosis. May be required for activation of the intrinsic but inhibit activation of the extrinsic apoptotic pathway. Acts as an antiapoptotic protein through association with GSK3A/B and BIRC2 in an apoptosis antagonizing signaling complex; activation of death receptors promotes caspase-dependent cleavage of BIRC2 and DDX3X and relieves the inhibition. May be involved in mitotic chromosome segregation. Is an allosteric activator of CSNK1E, it stimulates CSNK1E-mediated phosphorylation of DVL2 and is involved in the positive regulation of canonical Wnt signaling (By similarity).[UniProtKB/Swiss-Prot Function] |
Documents
| Product Manuals |
| FAQs |
| SDS |
Resources
Other Versions
| SKU | Description | Size | Price |
|---|---|---|---|
| KN304393BN | Ddx3x - mouse gene knockout kit via CRISPR, HDR mediated |
USD 1,657.00 |
|
| KN304393LP | Ddx3x - mouse gene knockout kit via CRISPR, HDR mediated |
USD 1,657.00 |
|
| KN304393RB | Ddx3x - mouse gene knockout kit via CRISPR, HDR mediated |
USD 1,657.00 |
|
| KN504393 | Ddx3x - KN2.0, Mouse gene knockout kit via CRISPR, non-homology mediated. |
USD 1,657.00 |
|
| GA201046 | Ddx3x CRISPRa kit - CRISPR gene activation of mouse DEAD/H (Asp-Glu-Ala-Asp/His) box polypeptide 3, X-linked |
USD 1,657.00 |
{0} Product Review(s)
Be the first one to submit a review